063-真核生物基因表达调控
刘小泽写于18.12.7
这个部分是现在研究的热门,因此很有必要了解一下背后的生物知识,我们不仅要做攻城狮,还要知道背后的生物意义,知己知彼,知根溯源,这样也不枉自己辛苦跑流程
首先看下真核生物的基因表达特点
- 基因组比原核生物大,且只有10%左右序列可以编码蛋白、rRNA、tRNA等(相比之下,原核生物大部分都是编码基因),其余的序列包括大量重复序列的功能目前还不清楚;
- 真核生物编码蛋白的基因不连续,转录后需要剪接去除内含子,这样就产生了可变剪切的分析,增加了表达调控的多样性;
- 真核生物中一个结构基因转录出一条mRNA,也就是说,mRNA是单顺反子(monocistron)【一个转录完毕的mRNA内含有外显子对应的转录产物,该mRNA只编码一条多肽链】。因此,下游产生的功能相关的蛋白或者一种蛋白的不同亚基将涉及多个基因的协调表达
- 真核生物核内基因与线粒体基因表达既独立又相互协调,核内基因与多种蛋白结合成染色质,直接影响基因表达
真核生物远比原核的表达调控复杂,真核的过程包括了:染色质激活、转录起始、转录后修饰、转录产物的细胞内转运、翻译起始、翻译后修饰等许多步骤。
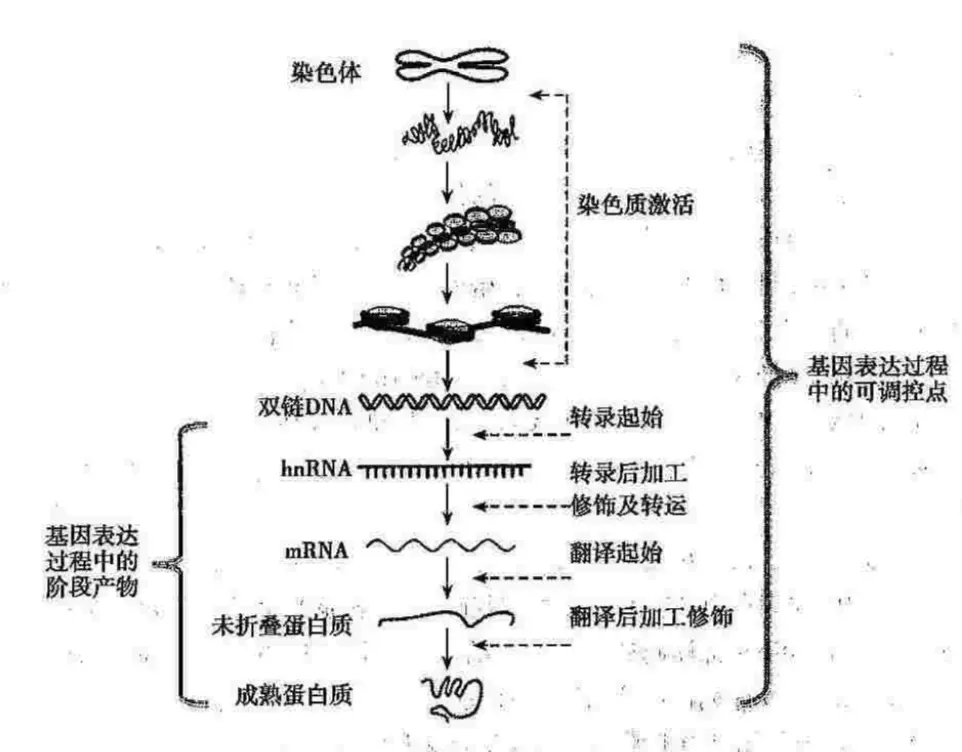
染色质激活
染色质的主要结构元件是核小体,核小体的核心区是4种组蛋白(H2A、H2B、H3、H4各2个分子)组成的八聚体,然后外面缠绕DNA双螺旋。其中每个组蛋白分子的氨基都会伸出核小体外 ,形成组蛋白尾巴【这些尾巴可以联系核小体,同时也是组蛋白修饰位点(修饰包括对组蛋白中富含的赖氨酸、精氨酸、组氨酸等带正电荷的碱性氨基酸进行的乙酰化、磷酸化和甲基化等过程)】
基因被激活时,染色质中核小体结构变得松弛,降低了核小体对DNA的亲和力,易于基因转录。
染色质水平控制基因转录的过程中,一个比较重要的机制就是DNA甲基化,这也是目前研究比较广泛的。甲基化简单说就是:胞嘧啶的第5位碳原子可以在DNA甲基转移酶(DNA methyltransferase)作用下被甲基化修饰为5-甲基胞嘧啶,常发生在CG序列中。另外,甲基化胞嘧啶在基因组的分布不均匀,有的地方含量比较高,一般将GC含量达60%,长度在300-3000bp的区段叫做CpG岛 。CpG岛主要在基因的启动子和第一外显子区域,大概超过60%的基因的启动子有CpG岛。染色质在转录活跃状态时,CpG岛的甲基化程度下降(如:管家基因的CpG岛中胞嘧啶甲基化水平较低)【CpG岛的高甲基化促进染色质形成致密结构,不利于基因表达】
染色质水平上的调控可以遗传给子细胞,原因就是细胞内存在具有维持甲基化作用的DNA甲基转移酶,可以在DNA复制后,按照亲本DNA链的甲基化位置对子链DNA的相同位置进行甲基化修饰【像这种遗传信息不在DNA中,而通过影响染色质结构及改变基因表达的方式,就是目前研究火热的**“表观遗传”(epigenetics)**】
转录起始
不管真核还是原核,转录起始都是基因表达调控的关键一步
对于真核生物来讲,必须知道一个东西:顺式作用元件 (cis-acting element)。它位于编码基因两侧,是可以影响自身基因表达活性的DNA序列,常常是非编码序列。真核基因组中的每一个基因都有自己特异的顺式作用元件,一般分为启动子、增强子、沉默子等
简单说,顺式作用元件就是:调控序列与被调控序列位于同一条DNA链; 我们还听过反式作用因子(trans-acting factor):调控序列远离被调控的编码序列,只能通过调控序列的表达产物发挥作用。这样的产物,不仅能对同一条链的结构基因进行表达调控,还可以对不在同一链的结构基因进行调控
启动子
一般包括转录起始点以及上游100-200bp的序列,包括许多功能独立的DNA序列元件,每个元件7-30bp左右。
众多的功能元件中,最经典的是TATA box,它的共有序列是TATAAAA,一般位于转录起始上游-25~-30bp区域,是基本转录因子TF II D的结合位点。除了TATA,还有GC box(GGGCGG)、CAAT box(GCCAAT)
增强子
长度大约200bp,顾名思义,就是增强转录效率(百倍以上)。核心组件常为8-12bp,单拷贝或多拷贝串联形式存在,另外也包含许多功能组件。增强子和启动子经常交错覆盖或连续排布。一般有以下的功能:
- 增强子与被调控基因位于同一条DNA链,属于顺式作用元件
- 增强子不仅可以在基因的上游或下游起作用,还可以远距离调节(通常为1-4kb),甚至更远可以影响到30kb以外的基因
- 发挥作用与序列方向无关,也就是说,将增强子方向倒置依然起作用【如果是启动子,倒置后就不能起作用】
- 增强子需要启动子才能发挥作用,但并不是说一个增强子就必须搭配一个启动子,它们之间是没有专一性的,同一个增强子可以影响不同类型的启动子
沉默子
属于负调控元件,作用不受序列方向影响,也能远距离起作用,可以对异源基因起作用
转录调控关键=》转录因子
转录因子(transcription factor, TF)是真核基因的转录调节蛋白。大多数的转录因子由其编码基因表达后,进入细胞核,然后识别、结合特异的顺式作用元件从而增强或降低相应基因的表达。
真核生物转录调控的基本方式:转录因子对顺式作用元件的识别与结合,通过DNA-蛋白的相互作用实施调控
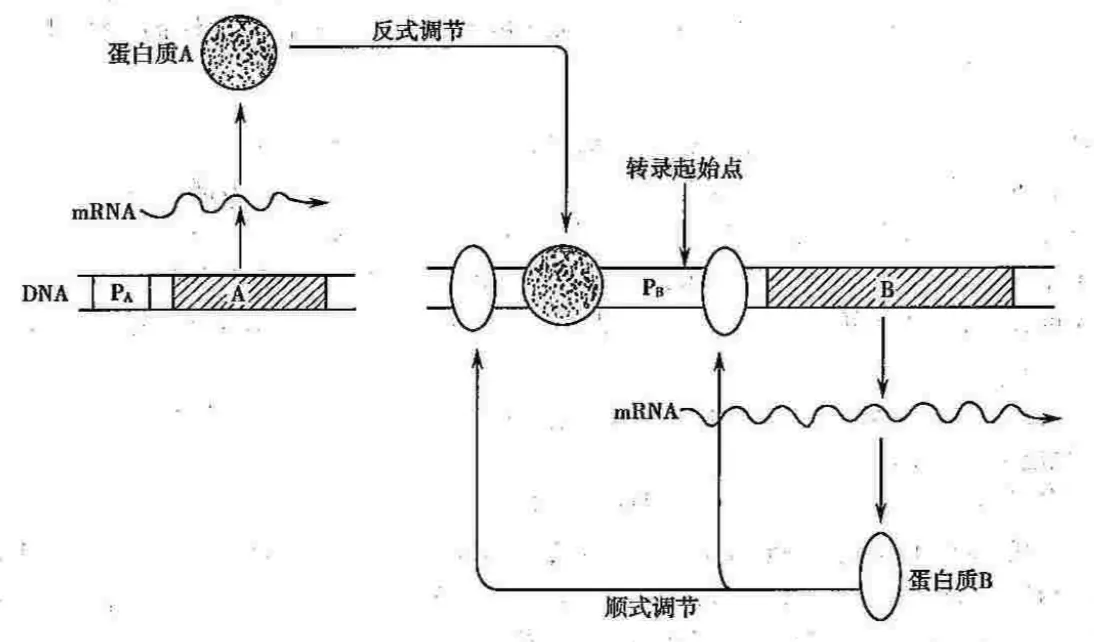
转录因子是反式作用因子;但是并非所有的转录调控蛋白都是反式作用,有些基因产物可以特异识别、结合自身的调节序列,这中调节蛋白就是顺式作用因子
上图中,蛋白A由它的编码基因表达后,通过与B基因特异的顺式作用元件的识别、结合,反式激活B基因的转录,因此蛋白A就是反式作用因子; B基因的产物可以特异性识别、结合自己的调节序列,B调节蛋白就是顺式作用因子
转录因子一般分为:通用转录因子和特异转录因子
通用转录因子
RNA聚合酶介导基因转录时必需的一类辅助蛋白,帮助聚合酶与启动子结合并起始转录【对所有基因都必需,没有组织特异性,因此也叫“基本转录因子”】
特异转录因子
个别基因转录需要,有的起转录激活作用(“转录激活因子”),有的转录抑制(“转录抑制因子”)。转录激活因子一般是一些增强子结合蛋白(enhancer binding protein, EBP);多数转录抑制因子是沉默子结合蛋白,当然还有不依赖DNA起作用的,比如通过蛋白-蛋白互作来“中和”转录激活因子,降低它们在细胞内的有效浓度,从而抑制基因转录。
关于结构:大多转录因子是DNA结合蛋白,至少有两个不同的结构域:DNA结合域(DNA binding domain)和转录激活域(activation domain)。其中DNA结合域主要有锌指模体(zinc finger)、碱性螺旋-环-螺旋(bHLH)、碱性亮氨酸拉链(bZIP);转录激活域分为:酸性激活结构域(与 TF II D协助)、谷氨酰胺富含结构域(与GC box结合)、脯氨酸富含结合域(与CAAT box结合)
转录后调控
因为真核生物的RNA产物要被送到细胞质中发挥作用,因此它的稳定性和降解过程影响了后续的基因表达
mRNA稳定性
在所有RNA分子中,mRNA寿命最短。真核生物的mRNA半衰期差别很大,长的有数十小时,短的只有几十分钟。一般半衰期短的mRNA多为编码调节蛋白,受环境变化大。总体上,影响mRNA稳定性的因素主要有:
- 5’帽子结构:作用是保护mRNA免除5‘-核酸外切酶的作用,并且与相应的帽子结合蛋白作用提高翻译效率,参与mRNA从细胞核向细胞质转运
- 3’polyA尾巴:作用是防止3‘核酸外切酶降解,还参与翻译起始过程
mRNA前体选择性剪切
转录产生的mRNA前体包含内含子、外显子。一般需要剔除内含子后变成一个成熟的mRNA,然后被翻译成多肽链。但实际上,参与拼接的外显子可以不按在基因组上的线性分布次序拼接,并且内含子也可以不完全切除。
选择性剪切的结果是**:同一条mRNA前体产生了不同的成熟mRNA**,并因此产生了不同的蛋白,这些蛋白的功能可以完全不同。
目前关于内含子的功能也是一个研究的方向,例如细菌丢失内含子,可以使染色体变小而加快复制速度;真核生物保留内含子,可以产生外显子移动,利于真核生物在环境改变时,合成功能不同但结构上仅有微小差别的蛋白
非编码RNA
- miRNA:属于小分子非编码RNA,长约22碱基;不同生物中有一定的保守性;有明显的表达阶段特异性和组织特异性;以单拷贝、多拷贝、基因簇等形式存在于基因组,大部分位于基因间隔区;通过碱基互补与特异的mRNA结合使其降解
- siRNA:干扰小RNA是一类双链RNA,可以通过酶切机制产生特定长度的(21-23碱基)和特定序列的小片段RNA。双链siRNA与特异靶mRNA完全互补结合,降解mRNA,组织翻译过程(即RNA干扰 RNA interference, RNA i)
- 长链非编码RNA(lncRNA):转录本超过200核苷酸,不直接参与基因编码和蛋白合成